8 overarching hypotheses – the basis of our research
H1: Both the persistence of the old associative trace and the acquisition of a novel inhibitory association characterize extinction learning as parallel events. In addition, a smaller part of the old trace is forgotten.
H2: Extinction procedures differ less on the behavioral level but still can vary substantially at the neural level. The specific biological salience and preparedness of CS and US affect excitatory and inhibitory learning, including extinction efficacy.
H3: The distinction between cues and context is learned. This learning requires the hippocampus because of its role in integrating stimuli over time and storing them as memories of past experiences. Prefrontal mechanisms interact with the hippocampus, also represent context and translate these context-dependent experiences into behavior.
H4: Sensory cortical fields and striatal territories are modulated by extinction learning. Thus, during extinction, activity patterns within both sensory cortical areas as well as corresponding dorsal striatal territories are altered.
H5: The cerebellum is part of the neural circuit underlying the different aspects of extinction, including context-related processes, conditioned fear and safety, and all other forms of associative learning. The cerebellum contributes to processing of prediction errors driving extinction learning, potentially within the time domain.
H6: Excitatory and inhibitory learning are phase-dependently affected by neuroendocrine and immune activation. These effects interact with stress-related emotional and cognitive alterations as well as with psychological comorbidities. Specific alterations in the extinction network are also relevant for learned immune responses.
H7: Understanding inter- and intra-individual variability in learning and extinction efficacy is crucial for translation. Impaired extinction contributes to pathology and/or to clinically relevant markers in healthy individuals.
H8: Avoidance and safety behaviors impact fear responses via different mechanisms than extinction learning. They counteract extinction learning efficacy by decreasing prediction errors that otherwise are key drivers of extinction learning.
The 8 hypotheses of the second funding period:
H1: Both learned inhibition and forgetting characterize extinction learning as parallel events.
H2: Appetitive and aversive extinction procedures differ less on the behavioral level but still can vary substantially at the neural level. The specific biological salience of CS and US affect excitatory and inhibitory learning, including extinction efficacy.
H3: The distinction between cues and context is learned. This learning requires the hippocampus be- cause of its role in storing memories of past experiences. Prefrontal mechanisms interact with the hippocampus to translate these context-dependent experiences into behavior.
H4: Sensory cortical fields show similar extinction learning properties as the BLA. These cortical fields will also modulate their striatal territories similarly as the BLA modulates its corresponding CEA segments. Thus, during extinction activity patterns within both sensory cortical areas as well as corresponding dorsal striatal territories are altered.
H5: The cerebellum is part of the neural circuit underlying the different aspects of extinction, including context-related processes, conditioned fear and safety, and all other forms of associative learning.
H6: Neuroendocrine and immune activation differentially affect extinction consolidation and its re- trieval. These effects are further modulated by the task-induced emotional arousal and context. The underlying mechanisms involve specific alterations in the extinction network and also apply to learned immune responses
H7: Understanding inter-individual variability and developmental changes in learning and extinction efficacy is crucial. Impaired extinction contributes to pathology and/or to clinically-relevant mark- ers in healthy individuals.
H8: Active avoidance impacts fear responses via different mechanisms than extinction learning. It counteracts extinction learning efficacy by decreasing prediction errors that otherwise are key drivers of extinction learning.
L1: The same model accounts for the dynamics of both acquisition and extinction in different learning paradigms and species. Different parameter settings in the model, such as learning rates, account for the variability across individuals, species, and paradigms.
L2: The trial-by-trial dynamics of behavior and psychophysiological variables increases (inhibitory learning) and decreases (forgetting) in associative strength during extinction.
L3: Context-dependence is learned, because the US is associated not only with the discrete CS, but also with diffuse contextual information.
N1: Functional and structural connectivity of the extinction network allow predicting inter-individual differences in the efficacy of extinction learning across paradigms (H7).
N2: Extinction of appetitive and aversive learning relies on partly distinct functional and structural connectivity patterns (H2).
N3: The cerebellum shows pronounced functional and structural connectivity with other areas of the extinction network. Connectivity patterns of different cerebellar subregions play specific roles for different aspects of extinction (H5).
N4: Functional and structural connectivity of the extinction network is systematically altered in pa- tients with disturbed extinction (phobia, chronic pain, cerebellar lesions; H7).
N5: Genetic variability predicts inter-individual differences of functional and structural connectivity of the extinction network.
The 7 hypotheses of the first funding period:
A more detailed overview of the 7 hypotheses from the first funding period can be found here. A technical description can be found here >>
1
Quick and easy:
During extinction learning, we not only forget the old memory but also actively suppress it.
Extinction learning involves both the deletion of the original association and the relearning of an inhibition:
this controversial assumption will be investigated in the SFB 1280 both using single cell recordings in animals, intracranial recordings in humans as well as and by conducting behavioral studies. During single cell recordings in animals with parallel optogenetic manipulation we expect (possibly in different brain regions) in the same experimental phase, some neurons stop responding to the conditioned, while other cell types may start to respond to the absence of the unconditioned stimulus. >>


Extinction learning of aversive and appetitive associations are manifested similarly in behaviour, but are partially distinct on a neurobiological level:
2
Quick and easy:
Whether we forget something bad or good seems completely the same to us in everyday life. However, different processes take place in our brain depending on the situation.
3
Quick and easy:
When we learn something, our environment plays a central role. Even if the context is not part of the actual learning process, it has a strong influence on it.
Context stimuli are only weakly associated with the unconditioned stimulus and control behaviour through the interaction of the prefrontal cortex and the hippocampus:
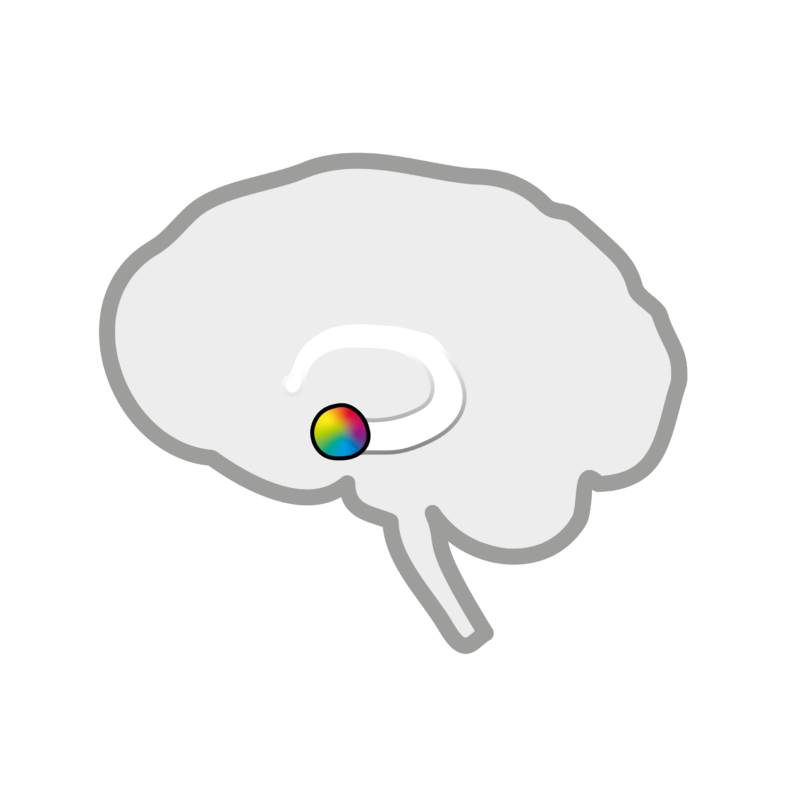
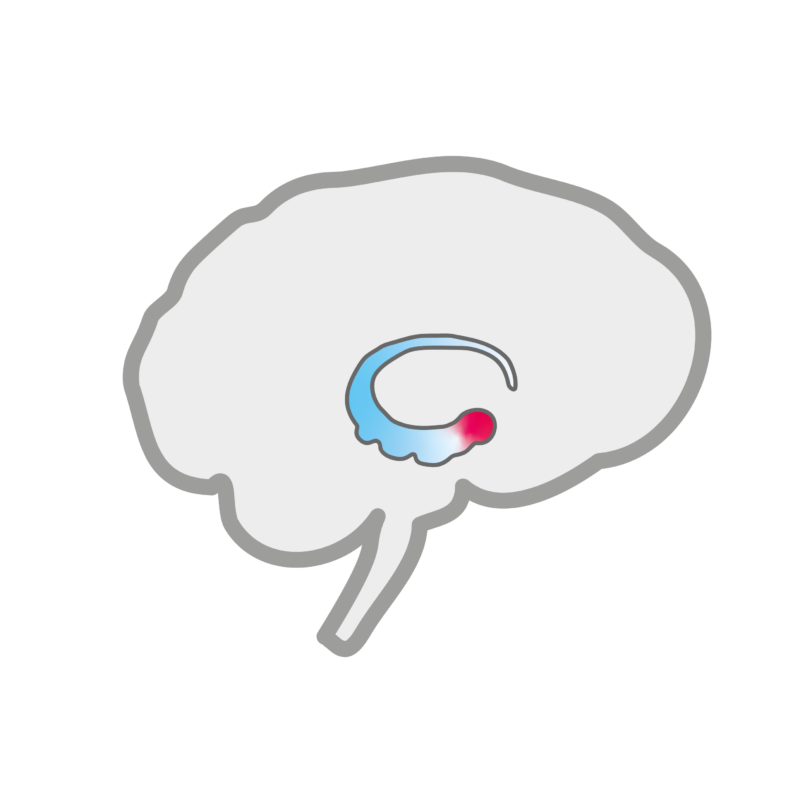
Schematic representation of the location of the amygdala in the brain.
The amygdala is a specialized corticostriatal system:
4
Quick and easy:
The amygdala is located deep in the brain and is known as our emotional center. New findings show that it is made up of several areas.
5
Quick and easy:
The amygdala,
together with our central control unit, the prefrontal cortex, and our memory center, the hippocampus, form the forgetting network in our brain. However, new findings show that other parts of the brain, such as the cerebellum (purple), also play a role in this process.
The neural extinction network is considerably larger than the triad of amygdala, hippocampus and prefrontal cortex:
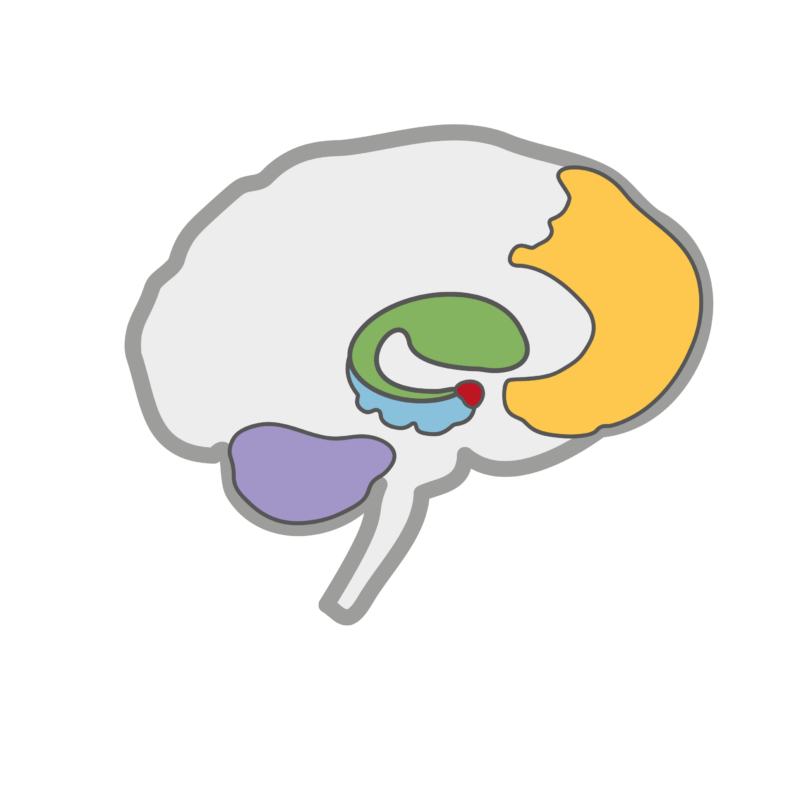
Long known as the extinction network: amygdala (red), hippocampus (blue) and PFC (yellow)

Central to the hormonal influence on our cognition:
The hypothalamus (pink) – pituitary glands (blue) – adrenal glands (green) – axis
6
Quick and easy:
Not only do our nerves (neurons) influence how we forget, our hormones are just as important for this process. Hormones and stress are closely linked. Anyone who has ever experienced a blackout due to high levels of excitement, for example, knows exactly how cortisol can affect our memory.
7
Quick and easy:
All of our brains are just as different as the rest of our bodies and different parts of the brain age faster than others. How these differences influence how we learn, forget and remember is what we want to find out in the SFB 1280.
Structure specific brain maturation as well as individual variability in extinction learning create different conditioning and pathological patterns:
Sources
Hadamitzky, M., Engler, H., and Schedlowski, M. (2013). Learned immunosuppression: extinction, renewal, and the challenge of reconsolidation. J. Neuroimmune Pharm. 8: 180–188.
Marsicano, G., Wotjak, C.T., Azad, S.C., Bisogno, T., Rammes, G., Cascio, M.G., Hermann, H., Tang, J., Hofmann, C., Zieglgänsberger, W., Di Marzo, V., and Lutz, B. (2002). The endogenous cannabinoid system controls extinction of aversive memories. Nature, 418: 530–534.
Medina, J.F., Nores, W.L., and Mauk, M.D. (2002). Inhibition of climbing fibres is a signal for the extinction of conditioned eyelid responses. Nature, 416: 330–333.
Swanson, L.W. and Petrovich, G.D. (1998). What is the amygdala? Trends Neurosci. 21: 323–331.
Wolf, O.T. (2017). Stress and Memory Retrieval: Mechanisms and Consequences. Curr. Opin. Behav. Sci. 14: 40–46.